MCA1とMCA2タンパク質が植物の接触センサーであることを特定
2021/10/20
- 研究
- プレスリリース
芝浦工業大学(東京都港区/学長 山田純)システム理工学部機械制御システム学科吉村建二郎教授、東京学芸大学(東京都小金井市/学長 國分充)飯田和子研究員、飯田秀利名誉教授の研究チームは、MCA1とMCA2タンパク質が、植物の接触センサーとしてはたらくことを世界で初めて証明しました。
植物に固有の接触センサーはいくつか候補があったにも関わらず、これまで特定されていませんでした。植物は強風・害虫・障害物などによる機械的刺激に遭遇しても移動できませんので、その場で対処するメカニズムを備えていると考えられています。本研究の成果はそのメカニズムの解明の糸口となり、さまざまな機械的ストレス下で植物が良好に成長する条件を突き止めることに繋がり、農作物の増産などに結び付くことが期待されます。
本研究の成果は2021年10月19日18:00日本時間で学術誌「Nature Communications」に掲載されます。
【芝浦工業大学ニュース】MCA1とMCA2タンパク質が植物の接触センサーであることを特定.pdf
 図1 指で触れられている植物
図1 指で触れられている植物
東京学芸大学の飯田秀利教授らは2007年に接触センサーの候補としてシロイヌナズナのMCA1とMCA2を発表しました(2)。MCA1とMCA2は互いによく似たタンパク質で、細胞膜に存在し、細胞膜を伸ばすなどの力学的刺激(=機械的刺激)を受けて細胞内にCa2+を取り込むはたらきをもっています。このようなはたらきをもつタンパク質を、専門用語でCa2+透過性機械受容チャネルと言います。
その後、シロイヌナズナのMCA1あるいはMCA2遺伝子を欠失している変異体の特徴を調べ、この遺伝子を酵母やアフリカツメガエルの卵母細胞内ではたらくようにしてMCA1とMCA2を解析することにより、両タンパク質はCa2+透過性機械受容チャネル(つまり、接触センサーの一種)の有力候補であることを示しました(2-4)。
しかし、決定的な証拠は当時得られませんでした。その理由は、MCA1とMCA2を発現させたのは生きている細胞なので、(a) MCA1とMCA2自身が接触や膜の伸展などの機械的刺激を直接感じているのか、それとも (b) 細胞内の別のタンパク質が機械的刺激を感じて、その情報をMCA1とMCA2に伝えているのかを区別することができなかったからです。(a)か(b)かを区別するには、精製したMCA1とMCA2タンパク質を脂質人工膜上に組み込んで、機械的刺激に応じたCa2+イオンの透過を電気生理学的に調べる必要があります。
また、研究チームは、MCA1とMCA2タンパク質は脂質人工膜上でそれぞれ4分子が会合してイオンチャネルを形成することを証明しました。この会合体は4本の茶筒が集まった形をしていると考えられます。その茶筒の真ん中に隙間ができます。この隙間をCa2+が通るというイメージです(図2)。ただし、このイメージどおりかどうかは今後の研究を待たなければなりません。
以上のように、芝浦工業大学と東京学芸大学の研究者が協力して、MCA1とMCA2が膜の伸展を直接感じてCa2+を透過させる機械受容チャネルであることを証明しました。図1で、植物の茎に指で触れるとその部分の細胞の膜が伸びます。その伸びを感じるMCA1とMCA2は接触センサーであると言えます。実際、MCA1とMCA2は茎だけでなく葉や根にも存在しますので、これらの器官で両者は接触センサーとしてはらたいていると考えられます。
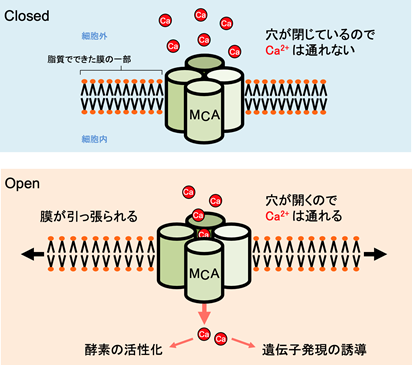 図2 MCA機械受容チャネルがCa2+を通す時の模式図
図2 MCA機械受容チャネルがCa2+を通す時の模式図
なお、MCA1とMCA2がCa2+を細胞内に流入させるはたらきがあるということは、両タンパク質は接触的刺激の最初の伝達体であるとも言えます。なぜなら、Ca2+は細胞外からの刺激を細胞内の酵素や遺伝子に伝える第二の伝達体(セカンドメッセンジャー)であることが知られているからです(図2)。
なお、動物のPiezoは接触センサーであることが証明されています。これを証明した研究者は米国のアーデム・パタポゥティアン(Ardem Patapoutian)教授であり、2021年のノーベル生理学・医学賞を受賞しました。一方、本研究は植物に固有の接触センサーを発見しましたので、今後学術的に高く評価されると考えられます。
植物への機械的刺激は、環境から加えられるだけでなく、細胞の分裂や伸長など自身の生命現象によっても加えられます。したがって、本研究の成果が植物の発生と成長のメカニズムの解明に貢献すると期待されます。
吉村建二郎 (芝浦工業大学システム理工学部機械制御システム学科教授)
飯田 和子 (東京学芸大学研究員、東京都医学総合研究所協力研究員)
飯田 秀利 (東京学芸大学研究員・名誉教授、元付属小金井小学校長)
論文名:
MCAs in Arabidopsis are Ca2+-permeable mechanosensitive channels inherently sensitive to membrane tension.
掲載誌:Nature Communications
DOI : 10.1038/s41467-021-26363-z
(10月19日18:00日本時間の解禁後にアクセス可能)
2)Nakagawa, Y., Katagiri, T., Shinozaki, K., Qi, Z., Tatsumi, H., Furuichi, T., Kishigami, A., Sokabe, M., Kojima, I., Sato, S., Kato, T., Tabata, S., Iida, K., Terashima, A., Nakano, M., Ikeda, M., Yamanaka, T., and Iida, H. (2007) Arabidopsis plasma membrane protein crucial for Ca2+ influx and touch sensing in rootProc. Nat. Acad. Sci. USA104, 3639-3644
3)Nakano, M., Iida, K., Nyunoya, H., and Iida, H. (2011) Determination of structural regions important for Ca2+ uptake activity in Arabidopsis MCA1 and MCA2 expressed in yeast. Plant Cell Physiol. 52, 1915-1930
4)Furuichi, T., Iida, H., Sokabe, M., and Tatsumi, H. (2012) Expression of arabidopsis MCA1 enhanced mechanosensitive channel activity in the Xenopus laevis oocyte plasma membrane. Plant Signal. Behav. 7, 1022-1026
5)Yoshimura, K., Usukura, J., and Sokabe, M. (2008) Gating-associated conformational changes in the mechanosensitive channel MscL. Proc. Nat. Acad. Sci. USA 105, 4033-4038
6)Yoshimura, K., and Sokabe, M. (2010) Mechanosensitivity of ion channels based on protein-lipid interactions. J. R. Soc. Interface 7, S307-S320
7)Iida, H. (2014) Mugifumi, a beneficial farm work of adding mechanical stress by treading to wheat and barley seedlings. Front. Plant Sci. 5:453. doi: 10.3389/fpls.2014.00453
植物に固有の接触センサーはいくつか候補があったにも関わらず、これまで特定されていませんでした。植物は強風・害虫・障害物などによる機械的刺激に遭遇しても移動できませんので、その場で対処するメカニズムを備えていると考えられています。本研究の成果はそのメカニズムの解明の糸口となり、さまざまな機械的ストレス下で植物が良好に成長する条件を突き止めることに繋がり、農作物の増産などに結び付くことが期待されます。
本研究の成果は2021年10月19日18:00日本時間で学術誌「Nature Communications」に掲載されます。
【芝浦工業大学ニュース】MCA1とMCA2タンパク質が植物の接触センサーであることを特定.pdf
ポイント
- MCA1とMCA2タンパク質が、植物の接触センサーとして働くことを世界で初めて証明
- 機械的刺激に遭遇した植物がその場で対処するメカニズムを解明する糸口となる
- 機械的ストレス下で植物が良好に成長する条件を突き止め、農作物の増産に期待
図1 指で触れられている植物研究の背景
オジギソウの葉は触られるとすぐに閉じることから、この植物は接触を感じられることが分かります。では、ほかの植物ではどうでしょうか(図1)。たとえば、花壇の植物の葉っぱに触っても目に見える反応はありません。しかし、実はどの植物も触られたことを感じているはずです。なぜなら、どの植物にも接触センサーがあると考えられるからです。進化論で有名なチャールズ・ダーウィンも1888年に出版した著書で、植物は接触を敏感に感じることができると書いています(1)。ただ、今日まで植物に固有の接触センサーの実体は特定されていませんでした。東京学芸大学の飯田秀利教授らは2007年に接触センサーの候補としてシロイヌナズナのMCA1とMCA2を発表しました(2)。MCA1とMCA2は互いによく似たタンパク質で、細胞膜に存在し、細胞膜を伸ばすなどの力学的刺激(=機械的刺激)を受けて細胞内にCa2+を取り込むはたらきをもっています。このようなはたらきをもつタンパク質を、専門用語でCa2+透過性機械受容チャネルと言います。
その後、シロイヌナズナのMCA1あるいはMCA2遺伝子を欠失している変異体の特徴を調べ、この遺伝子を酵母やアフリカツメガエルの卵母細胞内ではたらくようにしてMCA1とMCA2を解析することにより、両タンパク質はCa2+透過性機械受容チャネル(つまり、接触センサーの一種)の有力候補であることを示しました(2-4)。
しかし、決定的な証拠は当時得られませんでした。その理由は、MCA1とMCA2を発現させたのは生きている細胞なので、(a) MCA1とMCA2自身が接触や膜の伸展などの機械的刺激を直接感じているのか、それとも (b) 細胞内の別のタンパク質が機械的刺激を感じて、その情報をMCA1とMCA2に伝えているのかを区別することができなかったからです。(a)か(b)かを区別するには、精製したMCA1とMCA2タンパク質を脂質人工膜上に組み込んで、機械的刺激に応じたCa2+イオンの透過を電気生理学的に調べる必要があります。
研究の内容
芝浦工業大学の吉村建二郎教授は脂質人工膜を用いた電気生理学的研究において世界の第一人者です(5,6)。吉村教授は、東京学芸大学において試験管内で合成および精製されたMCA2を脂質人工膜上に組み込み、その膜を引っ張る(伸展させる)と、膜の伸展の度合いに応じて、MCA2がイオンを通す確率が上がることを発見しました。言い換えれば、MCA2は機械受容チャネルとしてはたらいたのです。また、MCA1またはMCA2を組み込んだ脂質人工膜の内腔にCa2+を結合する蛍光色素を入れておき、脂質人工膜内へのCa2+の流入を分光学的に調べました。その結果、MCA1またはMCA2を通して脂質人工膜内へのCa2+の流入が起こることも証明しました。また、研究チームは、MCA1とMCA2タンパク質は脂質人工膜上でそれぞれ4分子が会合してイオンチャネルを形成することを証明しました。この会合体は4本の茶筒が集まった形をしていると考えられます。その茶筒の真ん中に隙間ができます。この隙間をCa2+が通るというイメージです(図2)。ただし、このイメージどおりかどうかは今後の研究を待たなければなりません。
以上のように、芝浦工業大学と東京学芸大学の研究者が協力して、MCA1とMCA2が膜の伸展を直接感じてCa2+を透過させる機械受容チャネルであることを証明しました。図1で、植物の茎に指で触れるとその部分の細胞の膜が伸びます。その伸びを感じるMCA1とMCA2は接触センサーであると言えます。実際、MCA1とMCA2は茎だけでなく葉や根にも存在しますので、これらの器官で両者は接触センサーとしてはらたいていると考えられます。
図2 MCA機械受容チャネルがCa2+を通す時の模式図なお、MCA1とMCA2がCa2+を細胞内に流入させるはたらきがあるということは、両タンパク質は接触的刺激の最初の伝達体であるとも言えます。なぜなら、Ca2+は細胞外からの刺激を細胞内の酵素や遺伝子に伝える第二の伝達体(セカンドメッセンジャー)であることが知られているからです(図2)。
本発見の学術的位置付け
植物の接触センサーの候補にはMCA1とMCA2のほかに、Piezo、OSCA、MSL、TPKと呼ばれる機械受容チャネルがあります。このうち、MCA1とMCA2だけが植物に固有の機械受容チャネルで、その他の機械受容チャネルは動物または微生物にも類似のタンパク質があります。これまでの研究では、OSCAが機械的刺激を直に感じると証明されていました。本研究は、植物にのみ存在するMCA1とMCA2が接触センサーとしてはたらくことを世界で初めて証明しました。植物に固有の接触センサーを発見したことの意味は、植物だけにみられる機械的刺激への応答や植物体の成長のメカニズムを解明する手がかりを得たことです。この研究が進めば、やがて農産物の増収にもつながるものと期待されます。なお、動物のPiezoは接触センサーであることが証明されています。これを証明した研究者は米国のアーデム・パタポゥティアン(Ardem Patapoutian)教授であり、2021年のノーベル生理学・医学賞を受賞しました。一方、本研究は植物に固有の接触センサーを発見しましたので、今後学術的に高く評価されると考えられます。
本発見のインパクト
植物は生育に悪い環境でも移動することはできません。たとえば、強風、食害を起こす虫、伸長の妨げとなる障害物などに遭遇しても移動できませんので、その場で対処しなければなりません。そのため、植物は一般の人が思っている以上にこれらの機械的刺激を敏感に感じて対処しています。これらの感知と対処のメカニズムを知れば、植物の成長をより良く制御することができるはずです。この制御法を農業に適用すれば、農作物の増収に繋がるかもしれません。実際、日本では記録の残る限り江戸時代から機械的刺激を農業に利用してきました。それは麦踏みです。踏圧とも言います。秋まき麦の芽生えを冬に踏むことによって機械的刺激を与え、麦の増収につなげてきました(7)。植物への機械的刺激は、環境から加えられるだけでなく、細胞の分裂や伸長など自身の生命現象によっても加えられます。したがって、本研究の成果が植物の発生と成長のメカニズムの解明に貢献すると期待されます。
論文情報
著者 :吉村建二郎 (芝浦工業大学システム理工学部機械制御システム学科教授)
飯田 和子 (東京学芸大学研究員、東京都医学総合研究所協力研究員)
飯田 秀利 (東京学芸大学研究員・名誉教授、元付属小金井小学校長)
論文名:
MCAs in Arabidopsis are Ca2+-permeable mechanosensitive channels inherently sensitive to membrane tension.
掲載誌:Nature Communications
DOI : 10.1038/s41467-021-26363-z
(10月19日18:00日本時間の解禁後にアクセス可能)
参考文献
1)Darwin, C. (1888) The Movements and Habits of Climbing Plants. John Murray, London2)Nakagawa, Y., Katagiri, T., Shinozaki, K., Qi, Z., Tatsumi, H., Furuichi, T., Kishigami, A., Sokabe, M., Kojima, I., Sato, S., Kato, T., Tabata, S., Iida, K., Terashima, A., Nakano, M., Ikeda, M., Yamanaka, T., and Iida, H. (2007) Arabidopsis plasma membrane protein crucial for Ca2+ influx and touch sensing in rootProc. Nat. Acad. Sci. USA104, 3639-3644
3)Nakano, M., Iida, K., Nyunoya, H., and Iida, H. (2011) Determination of structural regions important for Ca2+ uptake activity in Arabidopsis MCA1 and MCA2 expressed in yeast. Plant Cell Physiol. 52, 1915-1930
4)Furuichi, T., Iida, H., Sokabe, M., and Tatsumi, H. (2012) Expression of arabidopsis MCA1 enhanced mechanosensitive channel activity in the Xenopus laevis oocyte plasma membrane. Plant Signal. Behav. 7, 1022-1026
5)Yoshimura, K., Usukura, J., and Sokabe, M. (2008) Gating-associated conformational changes in the mechanosensitive channel MscL. Proc. Nat. Acad. Sci. USA 105, 4033-4038
6)Yoshimura, K., and Sokabe, M. (2010) Mechanosensitivity of ion channels based on protein-lipid interactions. J. R. Soc. Interface 7, S307-S320
7)Iida, H. (2014) Mugifumi, a beneficial farm work of adding mechanical stress by treading to wheat and barley seedlings. Front. Plant Sci. 5:453. doi: 10.3389/fpls.2014.00453
お問い合わせ
芝浦工業大学 企画広報課
〒135-8548 東京都江東区豊洲3-7-5(豊洲キャンパス本部棟2階)
TEL:03-5859-7070 / FAX:03-5859-7071
E-mail:koho@ow.shibaura-it.ac.jp